
Motorisk kontrol
Motorisk kontrol er spørgsmålet om, hvilke principper der styrer de komplicerede bevægelser. Hvordan håndterer kroppen et system, der bl.a. består af hjernen, musklerne og leddene, og laver en koordineret bevægelse? Hvordan tager systemet højde for omgivelsernes omskiftelige karakter?
Disse spørgsmål forsøger jeg at besvare i denne artikel om motorisk læring, der blev bragt i Focus Idræt nr. 3 2007. Artiklen er baseret på mit speciale Kontrol og læring af motoriske færdigheder og verbal feedback, der er udgivet som bog bl.a. hos Saxo.

Kontrol og læring af motoriske færdigheder og verbal feedback
Bogen besvarer følgende to spørgsmål: (1) Hvordan kan komplicerede, hurtige færdigheder tænkes at være kontrolleret, og hvordan kan påvirkningen og læringen i dette kontrolsystem tænkes at foregå? (2) Hvordan kan underviseren verbalt give feedback på udøverens præstation af en kompliceret, hurtig færdighed for at skabe den bedst mulige motoriske læring hos udøveren?
Du kan læse hele artiklen herunder:
1 Indledning
At mennesket er i stand til at bevæge sig skyldes, at både reflekser og tillærte færdigheder spiller umærkeligt sammen. Når Ronaldinho går ind på fodboldbanen og pludselig mærker noget spidst i støvlen aktiveres hans automatiske bøjerefleks, og han trækker benet til sig for at beskytte foden. Dette er en medfødt refleks.
Når fodboldspillerne inden kampen står på rad og række på midten af banen og modtager publikums hyldest, bruger de stillingsrefleksen til at opretholde balancen. Denne refleks har stor betydning for kroppens holdning. De posturale muskler må hele tiden korrigere spillernes stilling, så de ikke falder bagover. Stillingsrefleksen er en tillært refleks.
Når kampen er fløjtet i gang, og Ronaldinho legende let snører den ene forsvarsspiller efter den anden, er hans tanker ikke optaget af alle de bevægelser, han render og laver. Hans tanker er formentlig mere optaget af strategien bag de handlinger, han gerne vil lave. En forsvarsspiller skubber til Ronaldinho, og han er ved at falde forover. Resolut genvinder han balancen ved at aktivere musklerne på bagsiden af underbenet og knæets bøjemuskler på lårets bagside, og han sparker på mål med et vristspark. Målmanden redder, og Ronaldinho går ud for at sparke hjørnespark. Det sparker han også med et vristspark. Begge færdigheder er tillærte, frivillige bevægelser, som han udfører uden bevidst indblanden, og da de har et startpunkt og et slutpunkt kan de karakteriseres som enkeltstående færdigheder. Efter lang tids træning er bevægelserne blevet automatiske.
Men færdighederne er også forskellige. Hjørnesparket kan karakteriseres som en lukket færdighed, fordi bolden ligger stille, og der er i princippet ikke noget, der forstyrrer udførelsen. Sparket på mål efter en dribletur kan kategoriseres som en åben færdighed, fordi Ronaldinho stilles over for en række uforudsigelige udfordringer; bolden ruller, modstanderne og medspillerne flytter sig, og målmanden agerer.
Retningsændringen og sparkene kan begge karakteriseres som hurtige bevægelser, fordi de udføres hurtigere end 200-300 ms. Det er nærliggende at antage, at kroppen kontrollerer færdigheder med varierende hastighed forskelligt. At flytte en skakbrik kræver opmærksomhed under hele udførelsesfasen, og det kan være nødvendigt at regulere bevægelserne undervejs. Det er usandsynligt, at afsættet i et længdespring eller et spark kan reguleres, når først det er sat i gang.
Selvom Ronaldinho ikke spekulerer over, hvordan han udfører bevægelserne, kan vi jo godt tænke lidt over det. Denne artikel opstiller en model for, hvordan komplicerede motoriske færdigheder, der udføres hurtigt, kontrolleres?
2 Motorisk kontrol
Motorisk kontrol og motorisk læring hænger uadskilleligt sammen. Studiet af motorisk kontrol er også studiet af, hvordan færdigheder læres gennem øvelse og træning (Singer, 1990). Men områderne kan analytisk behandles som to adskilte områder, fordi det først og fremmest er nødvendigt at vide, hvad der læres (altså et spørgsmål om motorisk kontrol), inden det er muligt at besvare, hvordan der skal læres (Whiting, 1992).
Motorisk kontrol er spørgsmålet om, hvilke principper der styrer de komplicerede bevægelser. Hvordan håndterer kroppen et system, der bl.a. består af hjernen, musklerne og leddene, og laver en koordineret bevægelse? Hvordan tager systemet højde for omgivelsernes omskiftelige karakter? Min indgangsvinkel til motorisk kontrol er inspireret af Agerskov (1994), som igen er inspireret af Bernstein (1984a), Arbib (1989) og Gallistel (1980). Motorisk kontrol er genstanden for denne artikel. I en senere artikel vil jeg forsøge at knytte motoriske kontrol og læring.
3 Teori om motorisk kontrol
I historien kan identificeres to paradigmer om motorisk kontrol (for gennemgange se Adams, 1987; Abernethy, 1992; Whiting, 1992; Summers, 2004). Jeg tager udgangspunkt i en teori udformet af den sovjetiske fysiolog Nikolai Bernstein og supplerer med Arbib (1989) og Gallistel (1980). Det gør jeg for det første, fordi teorien har haft stor indflydelse på opfattelsen af motorisk kontrol og læring. For det andet har teorien et hierarkisk udgangspunkt, fordi udførelsen af færdighederne påbegyndes i hjernen, men den ser også i høj grad kroppen som et selvorganiserende system, og det giver teorien en stor forklaringskraft. For det tredje er teorien funderet i et naturvidenskabeligt paradigme, men den har et stort forklaringspotentiale i forhold til den måde, man ofte taler om kroppen og bevægelser på i de paradigmer, der taler om kroppens tavse viden. For det fjerde kan teorien forklare kontrollen og læringen af både nye og eksisterende færdigheder, hvor andre teorier har haft vanskeligheder med nye bevægelsesmønstre (fx Schmidt, 1975). Og endelig for det femte tager teorien højde for eksterne kræfter og kan derfor forklare handlinger i tyngdekraftsfeltet.
3.1 Konstruktion af en færdighed
Motorisk kontrol kan analytisk opdeles i en forberedelsesfase og en udførelsesfase (Crow, 1998). I forberedelsesfasen beslutter kroppen hvilke bevægelser, der skal iværksættes på baggrund af stimuli både indefra og udefra. Forberedelsesfasen er en beslutningsproces, hvor udøveren vælger hvilket handlingssvar der er mest passende til situationen. Her fastsættes målet for udførelsesfasen.
Opbygningen af en frivillig handling (færdighed) kan beskrives af tre komponenter:
-
forsættet eller den motoriske opgave der skal løses. Opgaven formuleres i planlægningsfasen af bevægelsen og manifesteres neuralt i form af et motorisk program, hvilket jeg vender tilbage til.
-
et invariant resultat. Opgaven er konstant, og derfor skal resultatet også være konstant.
-
et varierende sæt af bevægelser. Bevægelser er aldrig metrisk i overensstemmelse med hinanden, fordi bevægelserne må tage højde for de skiftende kræfter i omgivelserne.
3.2 ‘Degree of freedom’-udfordringen
Bernstein (1984b) beskriver, hvordan en færdighed er et resultatet af koordinationen af et stort antal samarbejdende systemer. Motor cortex ikke blot er en fjernbetjening, der sætter forskellige bevægelser i gang. Frivillige, formålsrettede bevægelser skal også tage højde for kroppens nuværende stilling og position i rummet (Gurfinkel, 1998), og derfor kan det motoriske program ikke indeholde nøjagtige metriske forskrifter. Bernstein påpeger, at der ikke eksisterer en en-til-en overensstemmelse mellem de efferente impulser fra hjernen, og de bevægelser signalerne afstedkommer.
Der er i hvert fald fire forhold, der kan være med til at forklare, at der ikke er direkte sammenhæng mellem det igangsættende impuls og resultatet i periferien. For det første har vores led mange frihedsgrader. En færdighed kan udføres med varierende brug af de forskellige led. For det andet kræver en bevægelse, at en muskel får leddene til at bevæge sig. Systemet kompliceres af, at to ens sammentrækninger i en muskel kan have forskellig effekt alt efter sammentrækningerne i synergisterne og antagonisterne omkring musklen. For det tredje skal der være et system, der får musklen til at trække sig sammen med den rigtige hastighed og kraft på det rigtige tidspunkt. For at kunne udføre en færdighed korrekt, er det nødvendigt at de rigtige muskler kontraheres med den rette kraft på det rigtige tidspunkt. For det fjerde må vi tage højde for omgivelsernes skiftende eksterne kræfter og forskelle i initielle forhold.
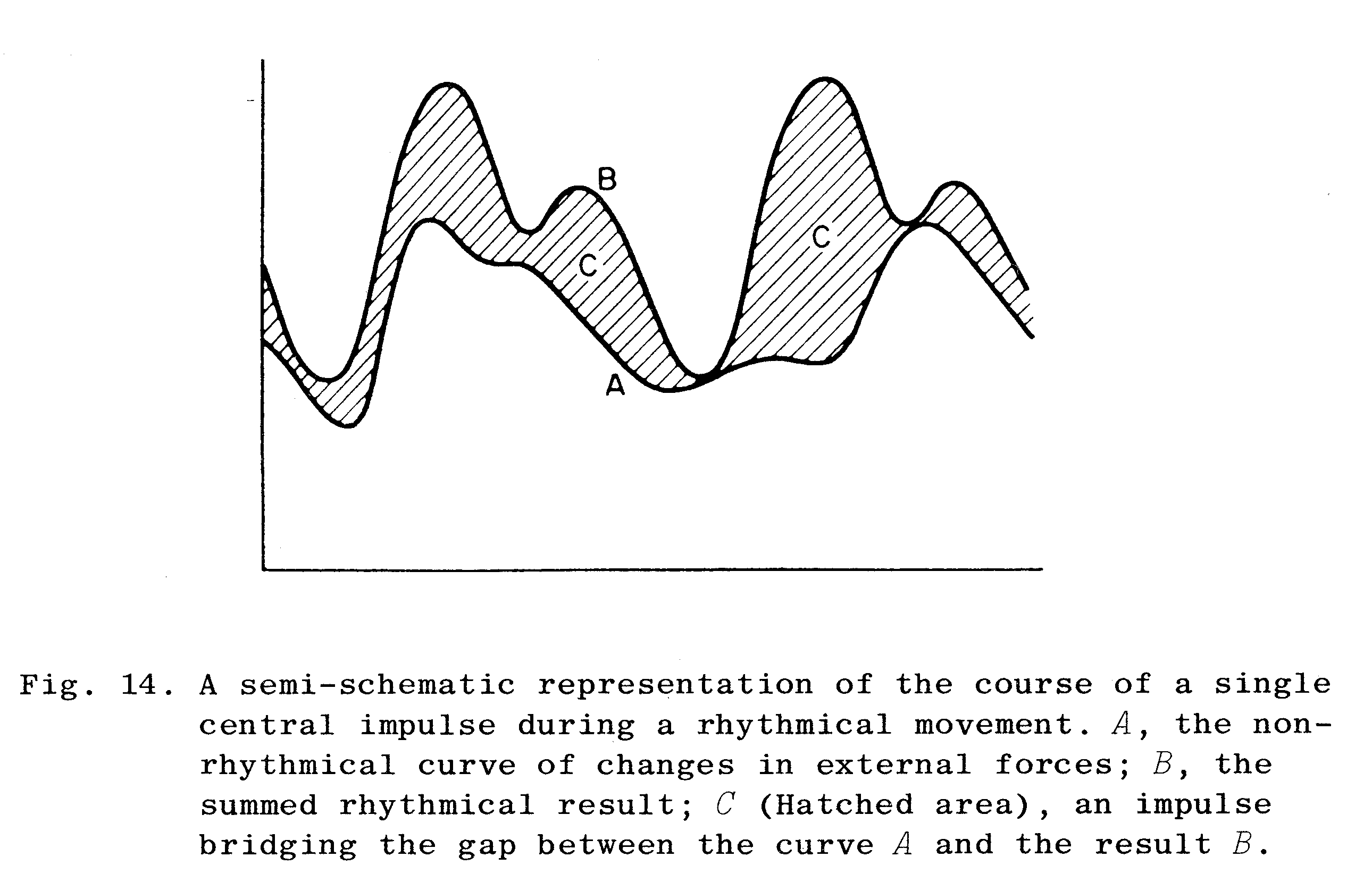
Figur 1 illustrerer det forhold der under en bevægelse eksisterer mellem den påkrævede kræftpåvirkning af et led (kurve B), de resulterende kraftpåvirkninger i omgivelserne (kurve A) og den supplerende kraft nervesystemet må bidrage med. Figuren viser, at der konstant sker ændringer i de kræfter som påvirker organismen. Det gør, at nervesystemet hele tiden må supplere med skiftende og passende kraftpåvirkninger indefra, så den resulterende kraftudvikling på leddene (kurve B) medfører den rigtige bevægelse (se også Agerskov 1994).
Den vigtigste biomekaniske forudsætning for at udføre en målrettet bevægelse er at overvinde det store antal frihedsgrader i kroppen og forvandle den til en kontrollerbar enhed. Nervesystemet står ved hver enkelt bevægelse over for det, Bernstein (1984a) kalder ‘degree of freedom problem’. Dette problem er for Bernstein det centrale problem, når man studerer motorisk kontrol.
3.3 Koordination og kontrol
Udøverens udfordring under frihedsgradsudfordringen er at koordinere bevægelsen af lemmerne, så et specifikt bevægelsesmål opnås. Bernstein (1984a) ser koordination som processen at mestre kroppens redundante frihedsgrader og derved gøre det muligt at kontrollere kroppen. Koordination er altså den proces der begrænser systemets frie variable og gør kroppen til en håndterbar organisering af kontrollen med det motoriske apparatet (Newell, 1984; Hodges, 2004).
Koordinationen kan enten handle om bevægelsen af en enkelt kropsdel eller om samspillet mellem flere kropsdele (Abernethy, 1992). For Bernstein er koordination et spørgsmål om at oprette nogle funktionelle muskel- og ledenheder. Men koordination skal også ses i forhold til de skiftende omstændigheder i omgivelserne (Bernstein, 1984b; Magill, 2001). Koordinationen foregår bl.a. gennem det motoriske hierarki (se mere senere) og selektiv potentiering (se mere senere).
Kontrol er den proces, som knytter værdier til variablerne i koordinationsfunktionen. Det er fx udsvinget, hastigheden, accelerationen og kraften i bevægelsen. Den velkontrollerede bevægelse er den, der bedst lever op til forventningen til bevægelsen. Motorisk kontrol er et spørgsmål om, hvordan kroppen løser et øjeblikkeligt motorisk problem skabt af omgivelserne (Whiting, 1992). For Bernstein er kontrol en omfattende integrering af flere systemer og systemets anticipation af, hvad der er nødvendigt for at kunne opnå målet med færdigheden. Derfor må bevægelseskonstruktionen tage højde for (Latash, 1998):
-
at organisere en cyklisk proces hvor feedback- og feedforward-sløjfer hele tiden spiller ind på bevægelsen.
-
at måle effektiviteten af kontrolstrategierne, hvilket sikkert er afgørende i færdighedstilegnelse.
Bernstein opdagede, at variabiliteten af bevægelsesbaner ikke er elimineret selv i højt automatiserede bevægelser. Bevægelserne bliver ikke identiske, selvom det motoriske resultatet bliver meget reproducerbart. Kun denne variabilitet sikrer den store præcision, når uventede kræfter spiller ind (Latash, 1998). Når tennisspilleren opfatter, at bolden tager et uventet opspring, så kan han korrigere slaget lidt, så han stadig får bolden over nettet. Slaget bliver måske ikke lige så hårdt og præcist, som det var meningen, men han formår at returnere bolden, selvom han var forberedt på noget andet (se også Agerskov 1994).
Variationer i én udførelsesvariabel bliver kompenseret for med tilpasninger i en anden; det fænomen som Bootsma kalder kompensatorisk variabilitet. Denne kompensatoriske variabilitet er med til at skabe det nogenlunde reproducerbare bevægelsesoutput (Whiting, 1992). Små korrektioner af bevægelser er et resultat af systemets dynamiske egenskaber, fx under skrivning hvor albuen flytter hånden, så den kan skrive det rigtige sted, og håndleddet laver finjusteringer for at kompensere i albuens placering og samtidig sørge for at slutproduktet (det skrevne) bliver rigtigt.
3.4 Hurtige bevægelser er forudprogrammerede
Bevægelsesreguleringen bygger på præprogrammerede kontrolprogrammer, som sender efferente impulser til systemet. Der er fire væsentlige argumenter for, at motorisk kontrol af hurtige færdigheder (af en varighed på maksimalt 200-300 ms) bygger på nogle præstrukturerede bevægelseskommandoer (Schmidt, 2000; Schmidt, 1997; Schmidt, 2003; Agerskov, 1994):
-
Behandling af sensorisk eller responsproduceret feedback er langsom, og mange hurtige færdigheder vil være overstået, inden tilrettelser på baggrund af feedback ville kunne effektueres.
-
Studier har vist, at hvis kompleksiteten af en færdighed stiger, så øges reaktionstiden også. Dette tyder på, at den komplekse færdighed tager længere tid at planlægge.
-
I studier hvor nervebanernes afferente input er afbrudt, har individet stadig kunnet kontrollere færdighederne.
-
Højt specialiserede udøvere har større neurale områder i hjernen end begyndere. Fx rapporterer Lüders (2001), at hjernebarkområdet, der repræsenterer venstre hånd, er størst hos eliteviolinister, og området er størst hos dem, der tidligst er begyndt at spille violin. Muligheden for at folk med de største hjernebarkområder bare har større mulighed for at blive gode, kan dog ikke udelukkes.
Resultaterne tyder på, at der er en grundlæggende motorisk repræsentation i hjernen af en færdighed. Hvordan kan færdighederne tænkes at være lagret i programmer? Hvad indeholder programmet? Hvilken rolle spiller afferent information i forbindelse med motorisk kontrol? Det er nogle af de spørgsmål, som vil blive besvaret i følgende gennemgang.
3.5 Færdighedernes invariante kendetegn
For Bernstein er det invariante i færdigheden et spørgsmål om målet med færdigheden og forventningen til opfyldelsen af færdigheden. At der er metriske overensstemmelser mellem lignende bevægelser er et spørgsmål om systemets dynamik og ikke en konsekvens af den centrale programmering, som fx (Schmidt, 1975) ville hævde.
Den oprindelige skemateori (Schmidt, 1975), som han har revideret (Schmidt, 2000) og selv kommenteret (Schmidt, 2003) postulerer en invariant timing i den motoriske repræsentation i hjernen på baggrund af et studium af W. J. Wadman, J. J. Denier van der Gon, R. H. Geuze og C. R. Mol fra 1979 (se Schmidt, 2003). Resultaterne viste, at aktivitetmønsteret i hhv. agonist og antagonist ikke blev påvirket af, om bevægelsen var blokeret eller ej. Dette indikerer, at et bagvedliggende motorisk program forårsager en invariant timing i aktiveringen af de enkelte motoriske enheder, og studiets resultater er en udfordring for Bernsteins teori.
Et motorisk program kan ifølge Bernstein imidlertid ikke indeholde informationer om, hvilke motoriske enheder der skal aktiveres af et aktionspotentiale på hvilket tidspunkt. Et motorisk program kan heller ikke indeholde oplysninger om musklernes kraftindsats, som var et element i den oprindelige skemateori (Schmidt, 1975), for hvis man laver lignende bevægelser i forskellige bevægelsesplaner (horisontalt eller vertikalt) virker tyngdekraften forskelligt på fleksor- og ekstensorgruppen. Når bevægelserne skal ligne hinanden, er det altså ikke muligt at skalere muskelkraften proportionelt.
3.6 Afferent input
For hvert bevægelsesforløb skal konstrueres et nyt sammenspil mellem interne og eksterne faktorer, fordi der ikke findes to fuldstændig ens situationer. Forudindstillingen for afsendelsen af de efferente effektoriske impuls må nødvendigvis bygge på afferente impulser (Bernstein, 1984b). Kun på den måde kan man sikre en glidende bevægelse, for udbredelsen af nerveimpulser tager tid, og en korrektur på nu-værdier ville ikke kunne udbredes hurtigt nok til at få indflydelse på den efterfølgende bevægelse (Agerskov, 1994).
Men bevægelser ligner også sig selv fra gang til gang. Det er sandsynligt at hver ny konstruktion bygger på tidligere konstruktioner, men de skal konstant tilpasses de skiftende kraftforhold i omgivelserne og internt i kroppen, fx skiftende træthedsmønstre og følelsestilstande.
I de hurtige, ballistiske bevægelser, hvor den motoriske kontrol kan beskrives som en open loop-mekanisme, er der ikke tid til korrigerende feedback under udførelsen af færdigheden.
Bernsteins teori er en teori om aktion og ikke reaktion. Alle sensoriske korrektioner må bygge på udøverens anticipation af de kraftpåvirkninger, der kræves for at få slaget til at lykkes, inden bevægelsen begynder. Det kræver anticipation af de eksterne kræfter (kurve A i figur 1) og af den nødvendige resulterende kraftpåvirkning over leddet (kurve B i figur 1 ). Justeringen af færdighederne bygger på afferent information fra proprioceptorer, trykreceptorer og synet. De afferente input spiller både en rolle ved dannelsen af det effektoriske output og evalueringen af bevægelsen (Agerskov, 1994; Latash, 1998). Disse indviklede sammenhænge er naturligvis også en årsagerne til at udførelse af færdigheder kan være vanskelige for begynderen, som endnu ikke kan anticipere omgivelserne. Agerskov (1994, side 24) giver et eksempel med udgangspunkt i forhåndsslaget i tennis:
I forhåndsslaget må spilleren ud fra boldens bane og hastighed og den ønskede placering af returneringen danne en efferent anticipation af de kraftpåvirkninger, der kræves for det ønskede slag. Det kræver en afferent information om den ønskede tilstand, en afferent information om den nuværende tilstand, en afferent information af påvirkende kræfter undervejs i slaget (…) og en afferent anticipation af feedbacken af den resulterende kraftpåvirkning over leddene (…).
Selvom der ikke er tid til korrigerende feedback under bevægelsesudførelsen, spiller feedback fortsat en rolle. Når bevægelsessekvensen initieres sendes et signal til hjernen, om hvad den kan forvente af effektorisk output og sensorisk input fra bevægelserne. Det ønskede bevægelsesforløb kalder Bernstein Sollwert, og det registrerede forløb kalder han Istwert. Sollwert er kroppens reference of correctness, der udformes når bevægelsen sættes i gang.
Begrebet ‘collorary discharge’ (Arbib, 1989) kan bruges til at uddybe, hvad Sollwert kan tænkes at være i praksis. Collorary discharge indeholder ifølge Arbib (1989) informationer om, hvad kroppen har brug for at få at vide, og hvilke sensoriske input den skal ignorere. Kroppen må vide, hvordan bevægelsen bør føles, og hvordan den rent faktisk føles for at kunne lære noget. Jeg kommer ind på feedback senere.
3.7 Motoriske skemaer
Bernstein har altså en opfattelse af, at der ligger en grundlæggende motorisk repræsentation i hjernen til grund for bevægelser. Han erkender, at den motoriske kontrol må tage højde for, at færdigheder varierer, men at de også ligner hinanden fra gang til gang, og dette kompromis udmønter sig i det Bernstein betegner som motoriske skemaer. De motoriske skemaer fastlægger de grundlæggende detaljer ved færdighederne - det der går igen - og er åben for variationer der sker fra gang til gang. Jeg bruger i den resterende del af artiklen motoriske skemaer, når jeg taler om det åbne og stabile mønster, der er en aktiv og dynamisk organisation af tidligere erfaringer.
3.8 Hvordan bruges skemaerne i praksis?
Det er interessant, hvordan skemaerne bruges i praksis? Arbib (1989) skelner mellem en master kopi og en aktiv kopi af det motoriske skema. Denne opdeling gør det muligt at finde den ønskede færdighed, men samtidig gives en teoretisk løsning på, hvordan skemaet kan tilpasses situationen. For at aktivere et specifikt skema skal der være nogle informationer i omgivelserne, der danner basis for udvælgelsen. Den aktive motoriske master danner baggrund for indsamling af de sensoriske input, der har betydning for færdigheden. Men skemaet må også styre hvilke sensorisk informationer, der indsamles undervejs i bevægelsen (feedforward) og specificere hvad der er acceptable afvigelser fra idealbevægelsen (se også Agerskov, 1994).
Arbib (1989) hævder, at et individ også har et perceptionsskema som styres af den aktuelle opgave. Feigenberg (1998) påpeger på baggrund af en række forsøg med reaktionstid, at en probalistisk prognose foregår både sensorisk og motorisk. Individet anticiperer altså fremtiden med sanserne, og musklerne forbereder sig også. Han konkluderer, at reaktionstiden afhænger af, hvor dygtigt man forudser fremtiden (den probalistiske prognose) og af signifikansen af et givent stimuli. I reaktionsforsøget viste det sig, at selvom der var størst sandsynlighed for at, deltageren skulle reagere på et givent lyssignal, så kunne deltageren opnå samme reaktionstid på et andet lyssignal, der havde større signifikans for deltageren, fordi deltageren får en belønning for at reagere hurtigt på det alternative lyssignal (Feigenberg, 1998). Den probalistiske prognose baserer sig på tidligere erfaringer, og den danner baggrund for, hvordan programmet udformes for at kunne opfylde den ønskede fremtid.
Organismen er ikke bare en passiv modtager af sensorisk input, men den bearbejder det aktivt. Omverdenen er styrende for hvilket skema, der udvælges, men skemaet er også styrende for hvilke input individet søger efter. Dette er illustreret i figur 2. Det er sandsynligt, at individets undersøgelse af omverdenen er målrettet af anticipatoriske skemaer.
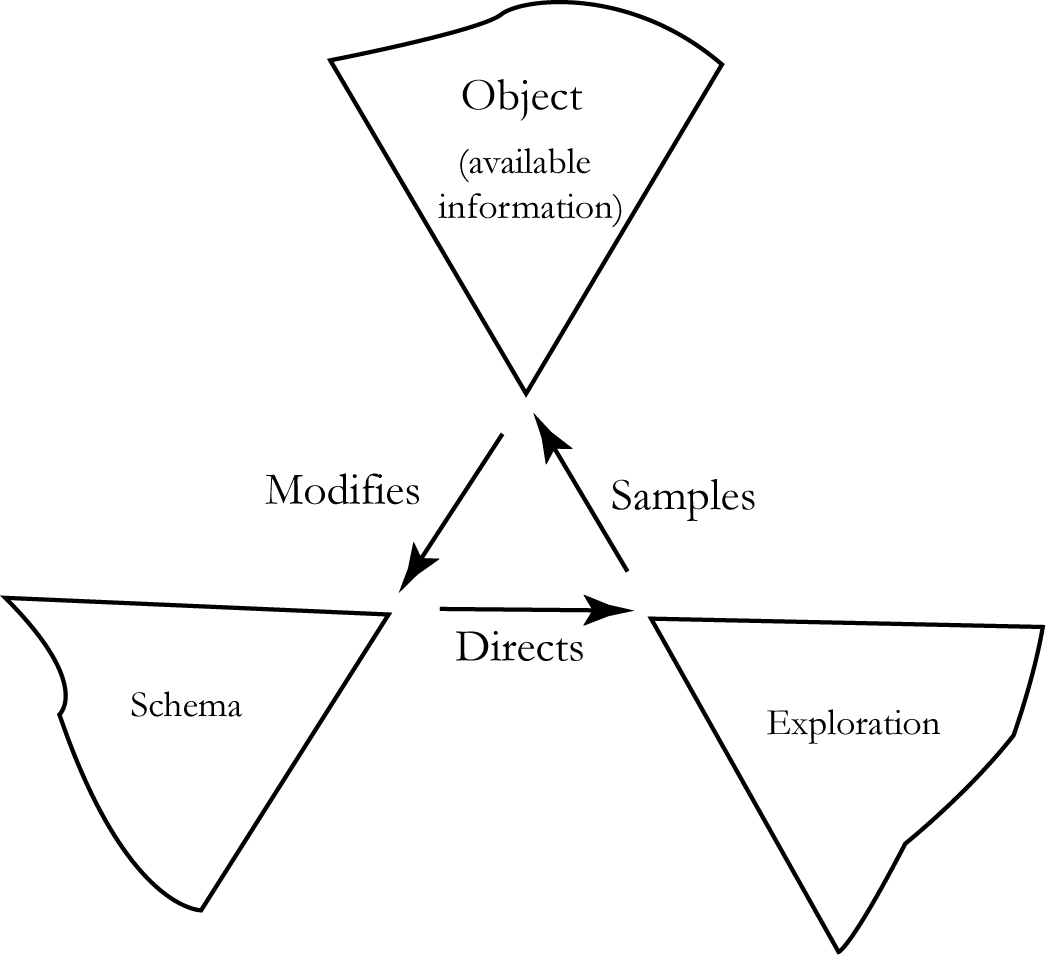
Kontrollen med en færdighed kan sandsynligvis overlades til ikke-bevidste kontrolområder, når den er højt automatiseret. Det sker fordi individets anticipatoriske skemaer er så veludviklede, at individet nøjagtig ved, hvad vedkommende skal være opmærksom på og derfor sorteres de sensoriske input hurtigere.
3.9 Topologisk repræsentation
Der er ikke en-til-en sammenhæng mellem efferente signaler og bevægelsesoutputtet. De afferente signaler fra musklerne siger imidlertid ikke noget om, hvordan bevægelsen forløber. Altså kan færdigheden ikke overordnet være kontrolleret af hverken efferente eller afferente signaler (Agerskov, 1994). Skemaet må være repræsenteret på anden vis.
Bernsteins tese er, at der på højere niveauer af centralnervesystemet eksisterer projektioner af rum og ikke projektioner af led og muskler. Færdigheden er planlagt med henblik på sit mål og ikke ned til den mindste detalje, hvor alle muskelkontraktioner er beskrevet (Weisendanger, 1998).
Bernstein mener, at færdighederne er beskrevet topologisk i hjernen, snarere end metrisk. Med topologisk mener Bernstein den spatio-temporale geometri, mens han med metrisk mener målinger af amplitude og hastighed. Færdigheden er altså gemt i et mentalt billede. Topologien er specificeret vha. den spatielle bane eller som et målrettet slutpunkt. Bernstein finder bl.a. et indicium for denne sammenhæng ved skrivning, hvor der uanset størrelsen af bogstaverne er særlige kendetegn, som kan tilskrives den topologiske beskrivelse af færdigheden i hjernen. Bernstein introducerer princippet om ‘equal simplicity’, som illustreres i figur 3. Figuren illustrerer en person der i luften tegner to cirkler, der ligner hinanden. Den ene tegnes umiddelbart foran vedkommende, og den anden tegnes lateralt i forhold til kroppen. Hans argument er, at færdigheden ikke kan være styret af et skema, der indeholder beskrivelser af muskelinnervationen, da de to bevægelser har forskelligt musklinnervationsmønstre. Alligevel udføres det med omtrent samme lethed og præcision (Weisendanger, 1998). Bevægelserne virker til at være styret af den samme motoriske repræsentation, men bevægelserne forårsages af forskellige ledbevægelser og muskler.
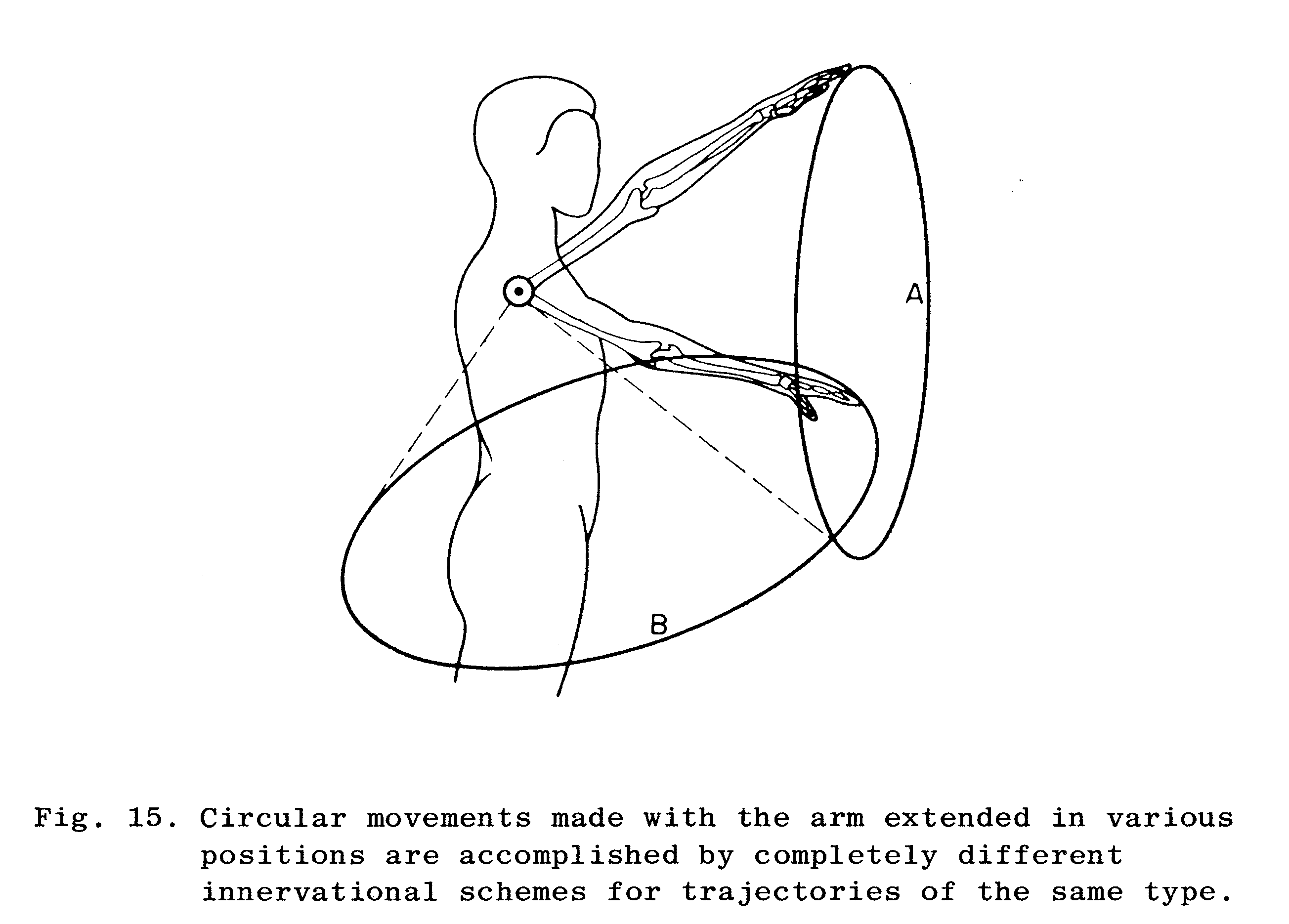
Hvis alle aspekter af den motoriske kontrol er defineret på højeste niveau ville det kræve en ekstrem stor processeringskapacitet, og efter al sandsynlighed ville den komme til at foregå temmelig langsomt. Derfor giver det mening, at den besluttende part ikke specificerer hele færdigheden ned til motorneuron-niveau; den centrale repræsentation er ikke specifikke kommandoer til de enkelte muskler.
Det er en stor opgave at få sendt de rigtige efferente impulser af sted på netop de tidspunkter, hvor musklerne ikke befinder sig i en refraktærperiode. Et efferent signal løber efter alt-eller-intet-princippet. Bevægelserne må være koordineret forud for de effektoriske impulser. Det er således ikke den centrale impuls, som alene bestemmer den motoriske effekt i periferien. Den motoriske effekt er et resultat af mange forskellige afferente signalers påvirkning af den centrale impuls (Agerskov, 1994).
Det indebærer, at kontrol i hvert fald foregår på to niveauer:
-
et øvre niveau, hvor færdigheden målrettes og der fastlægges en topologisk beskrivelse af færdigheden.
-
og et nedre niveau som er ansvarlig for den metriske udførelse.
Kernen i teorien er stabilitet, som kroppen vil forsøge at opnå, hvis systemet er forstyrret. Stabiliteten er kendetegnet ved, at variabiliteten er minimeret i bevægelserne, og færdigheden er udført med de mest energiøkonomiske bevægelser (Magill, 2001). Kroppen er en selvorganiserende enhed, hvor den motoriske kontrol er distribueret, og systemet finder en løsning på en given bevægelsesudfordring.
Med disse overvejelser kan jeg yderligere præcisere, at koordination for en given færdighed kan siges at være de topologiske karakteristika for en relativ færdighed, mens kontrol er skaleringen eller de absolutte værdier, der knyttes til en færdighed (Newell, 1984).
3.10 Kinæstetisk repræsentation
Agerskov (1994) tillægger imidlertid også den kinæstetiske repræsentation betydning for den motoriske kontrol. Den kinæstetiske repræsentation supplerer den topologiske repræsentation. Fx vil tennisspilleren forsøge at komme i den rigtige udgangsposition inden udførelsen af et slag i tennis. Spilleren vil forsøge at minimere variationen i de eksterne kræfter så meget som muligt. Hvis udøveren formår at skabe denne ensartethed i bevægelsessituationen og derved opnå den samme fornemmelse hver gang færdigheden udføres, så begynder “ muskeltonus at kunne bruges som information om bevægelsesudførelsen”.
Det motoriske skema kan både findes som et topologisk skema og et kinæstetisk skema. “Fornemmelsen” er ikke bare et abstrakt, fænomenologisk begreb. Udøveren har helt konkret lært hvilke afferente sensoriske effekter en automatiseret færdighed har, men også hvilke sensoriske input der almindeligvis går forud for en bevægelse. Bernsteins topologiske skema bør altså i det mindste suppleres med et kinæstetisk skema.
3.11 Kludder i udvælgelsen af skemaer og emotionel repræsentation
Bernsteins teori er fysiologisk i sit sigte og beskæftiger sig ikke med psykologiske aspekter. Den bruger mest tid på, hvordan kontrollen er i udførelsesfasen af færdigheden. Under planlægningskontrollen mener jeg dog, at det kan være sandsynligt, at et emotionelt aspekt også kan spille ind. Ligesom der gemmes et kinæstetisk skema er det sandsynligt, at der også i den episodiske hukommelse gemmes et emotionelt skema i forbindelse med en færdighed (Selch, 2003).
Dette forhold kan illustreres med et golfsving. Udøveren stiller op og gør klar til at slå. Han kan trække på det topologiske skema for færdigheden, og han ved, hvordan udførelsen af færdigheden bør føles. Han slår alligevel bolden direkte ud i roughen. Han har sit topologiske skema på plads, og bevægelserne giver ham nogle kinæstetiske informationer. Ud over det får han imidlertid også en følelse knyttet til golfsvinget. Formentlig en følelse af ærgrelse. Han stiller sig op igen. Han kan stadig trække på det topologiske og kinæstetiske skema, men det emotionelle skema er nu også til stede. Måske får det emotionelle skema ham til at tvivle på det kinæstetiske og i sidste ende det topologiske skema.
Der findes teorier om, at man hurtigt kan fremdrage begivenheder og episoder knyttet til følelser. På samme måde kan man opstille en hypotese om, at skemaer der har knyttet en emotionel repræsentation til sig, bliver domineret af den emotionelle repræsentation.
3.12 Men hvordan kommer vi fra skema til bevægelse?
Hvis motoriske skemaer er en aktiv organisering af tidligere erfaringer og den enhed hvorfra den motoriske kontrol koordineres, så er det interessante spørgsmål, hvordan koordineringen foregår?
Færdigheden er repræsenteret topologisk og kinæstetisk (og måske emotionelt) i hjernen; det er et skema, som ikke indeholder nogen information om muskelinnervation. Det er en kommando om en bestemt færdighed, som sendes ud fra centralnervesystemet. Formålet med det følgende er at beskrive, hvordan skemaerne kan forbindes med den konkrete muskelinnervationen.
Skemaer er et neurologisk funderet mønster, som bygger på tidligere erfaringer. Det er vanskeligt helt nøjagtigt at kortlægge et skema, fordi det ikke følger en udbredelsesbane fra neuron til neuron, men det er snarere et mønster, som gør systemet i stand til bedre at kunne håndtere kommende situationer (Agerskov, 1994). Hypotesen er, at nervesystemet hver gang det reagerer korrekt, styrker det anvendte skema, og hvis der ikke er et dækkende skema til stede, vil nervesystemet justere på mønstrene for næste gang at kunne aktivere et skema, der er mere hensigtsmæssigt til at løse den situationsbestemte motoriske opgave. Systemet bliver selvorganiserende, når det skal give løsninger til motoriske problemer. De mange frihedsgrader er reduceret til nogle enkelte kontrolparametre i et håndterbart system (Whiting, 1992).
Hjernen formår i læringsprocessen at fastholde erfaringerne i et mønster, der fastholder det gentagne og er åben for det varierende. Hvis hele færdigheden skulle specificeres i hjernen, ville det kræve en rigtig stor processeringskraft, når kroppen skal tage højde for alle frihedsgraderne og de eksterne kræfter.
Problemet løses med det motoriske hierarki, og det følgende afsnit besvarer, hvordan hele færdigheden organiseres på flere niveauer i kroppen?
3.13 Motoriske hierarki
I det motoriske hierarki fordeles kontrollen på flere niveauer. Bernstein opererer med tre handlingssystemer. Den primære opgave for det laveste niveau (A) er at opretholde balancen. Dette niveau er praktisk talt ikke tilgængeligt for bevidstheden. Niveau B er nødvendigt for bevægelse og gentagne, rytmiske bevægelser, og kun semi-voluntære handlinger er kontrolleret på dette niveau. Niveauet for rummet (C) er ansvarlig for målrettede bevægelser, og på dette niveau er processeringen af information kritisk. Det må altså være på dette niveau arbejdshukommelsen og den topologiske repræsentation er til stede. Det interessante er, at integreringen af niveauerne skifter alt efter færdighederne eller udøverens niveau. Bernsteins motoriske hierarki tager tydeligt udgangspunkt i den selvorganiserende krop, men det er ikke tydeligt, hvor muskler og led hører hjemme.
Det får vi et klarere billede af hos Gallistel (1980), som ser bevægelseskoordination som værende organiseret i et hierarki med seks niveauer; set nedefra starter det med de motoriske neuroner og set ovenfra starter det med organismen som et hele.
- Den motoriske enhed.
- En muskel (fx mm. quadriceps femoris), der udgør koordinationen af de motoriske enheder.
- Muskelgruppe (fx en fleksorgruppe ved knæleddet).
- Organniveau (fx koordinationen af et ben).
- Organsystem (fx begge ben under gang og løb).
- Kroppen som helhed.
Hierarkiet er udformet så enhederne på de forskellige niveauer er relativt autonome. Hvert niveau opererer selvstændigt ud fra ordrer fra et højere niveau. Den øverste enhed bestemmer ikke, hvad der skal ske på det laveste niveau. På det øverste niveau 6 tages beslutningen om hvilken færdighed, der skal udføres. Det er her planlægningen og programmeringen foregår (svarer til Bernsteins niveau C). Derefter går beskeden nedad i systemet, men systemet organiserer sig selv. Indflydelsen oppefra består i at sænke innervationstærsklen hos nogle enheder og hæve den hos andre; nogle enheder får tilladelse til at blive aktiveret, mens andre ikke får det (Gallistel, 1980). De lavere niveauer har sensorisk indflydelse på, hvordan en given handling udføres, eller i praksis den metriske udformning, mens de højere niveauer planlægger handlingen gennem det topologiske skema.
En motorisk respons er styret af det motoriske problem og det ønskede motoriske resultat. Det er to invariante størrelser. Målopnåelsen sker vha. variante processer. Det er i Gallistels model niveau 6, der sammensætter bevægelserne, så målet for færdigheden opnås. De motoriske skemaer findes på niveau 6, som kan siges at være mødestedet mellem image of the act og de effektoriske signaler, der skal igangsætte udførelsen af færdigheden, og det er også på niveau 6 motivation og kognition findes (Agerskov, 1994).
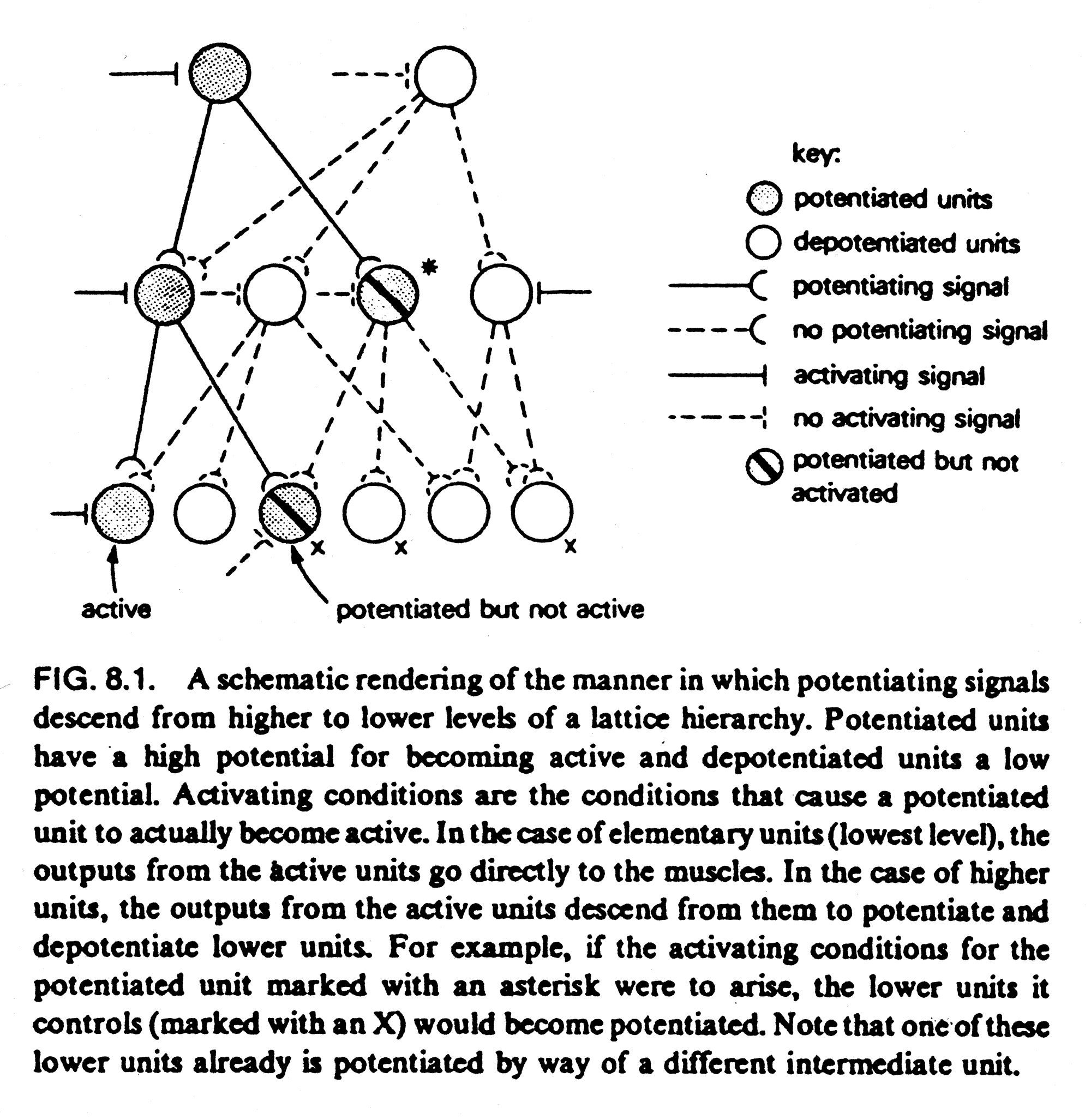
De enkelte autonome enheder agerer naturligvis ikke fuldstændig uafhængigt af hinanden. Hvert niveau kan derimod “operere som autonome processer på basis af parametre dikteret fra højereliggende niveauer” (Agerskov, 1994, side 41). Det motoriske hierarki sender ikke bare signaler fra de højere niveauer, som serielt aktiverer neuroner længere nede i systemet og til sidst aktiverer de motoriske neuroner, der aktiverer et aktionspotentiale og derved en sammentrækning af de motoriske enheder. Aktiveringen foregår i stedet gennem et distribueret mønster i det Gallistel kalder et latticehierarki (figur 4), hvor de overliggende niveauer påvirker de underliggende niveauer ved at potentiere og depotentiere forbindelserne i hierarkiet. De underliggende niveauer tillades kun at være aktive inden for visse rammer og grænser for, hvordan det er mest hensigtsmæssigt at reagere i den pågældende situation. Denne selektive potentiering giver altså systemet mulighed for lave en bevægelse. Bevægelsesreguleringen er selvregulerende på alle niveauer inden for grænserne specificeret af niveauet højere.
Hvis man fx aktiverer kattens bøjerefleks i svingfasen med en berøring af poten, vil der ske en øget fleksion i benet. Hvis samme berøring sker i standfasen vil katten i stedet lave en øget ekstension for at komme hurtigere væk fra stedet (Gallistel, 1980; Agerskov, 1994). Selektiv potentiering er et meget afgørende princip i latticehierarkiet.
Figur 4 viser, at de underliggende niveauer kan have flere end et overordnet niveau. Det kalder Gallistel ‘common path principle’. Det betyder, at de underliggende niveauer kan indgå i flere sammenhænge. Common path og selektiv potentiering betyder også, at det samme stimulus kan aktivere forskellige reaktioner.
Lad os igen tage et eksempel fra en tennisserv. Spilleren vælger at ville server udadskruet og dybt i banen. Han vælger det relevante topologiske skema. Den aktive kopi af skemaet udvalgt på niveau 6 sætter parametrene for niveau 5, hvor benenes og armenes indbyrdes koordination organiseres. Fra organsystemet skal timingen af benets bøjefase, kroppens centralbevægelse og armens svingfase koordineres på niveau 4 gennem en temporal bestemmelse af innervationstærskler fra niveau 5. Når denne timing er på plads sørger niveau 4 for retningslinjer til muskelgrupperne på niveau 3. Hvert niveau fungerer autonomt inden for de overordnede grænser. Hvis opkastet af bolden bliver for lavt, skal armens svingfase måske justeres i forhold til boldens bane, eller også skal spilleren stoppe udførelsen af bevægelserne, lade bolden falde og forsøge igen.
Det enkelte niveau kan kun fungere som en autonom enhed, hvis der er en sammenhæng mellem efferente og afferente signaler. Hvis et niveau udelukkende fungerede ud fra et effektorisk bevægelseskommando, ville der ikke være nogen reguleringsfunktion på niveauet. Den afferente dimension, som Bernstein lægger stor vægt på, mangler i Gallistels hierarki (Agerskov, 1994). Uden en feedforward-mekanisme ville der kun være mulighed for anticipation på det overordnede niveau 6.
Hvis systemet skal kunne tilpasse sig, er der nødt til at være anticipation på hvert niveau af hierarkiet, og det kræver at der både er feedforward og feedback på alle niveauer. Anticipationen fungerer naturligvis forskelligt på de forskellige niveauer. På niveau 6 er anticipationen en abstrakt ide om færdigheden - altså en topologisk repræsentation af færdigheden. På de underliggende niveauer eksisterer anticipationen som ubevidste neuromuskulære processer.
3.14 Behandling af feedback
Det motoriske hierarki har brug for feedback. De forskellige receptorer rundt i kroppen registrerer kroppens aktuelle tilstand og sender beskeder gennem nervetrådene. Disse feedbackløkker er dog forsinket i større eller mindre grad (tabel 1), sandsynligvis efter hvor langt feedbacken skal vandre i det motoriske hierarki, hvilket komplicerer systemets brug af feedback, fordi den interne feedback har en latenstid på mellem 30 og 180 ms.
[Table 1: Tidsrummet fra et feedback-loop er sat i gang til det når bestemmelsesstedet (Schmidt, 2000)]
| Latency | Flexibility | Role of instructions | Effect of number of choises | ||
|---|---|---|---|---|---|
| M1 response | 30-50 | Almost none | None | None | |
| M2 response | 50-80 | Low | Some | None (?) | |
| Triggered reaction time | 80-120 | Moderate | Large | Moderate | |
| Reaction-time response | 120-180 | Very High | Very large | Large |
M1-refleksen er en monosynaptisk refleks, som kun går via en synapse mellem den sensoriske nerve og direkte hen til den motoriske neuron. Et eksempel er knæstrækkerrefleksen.
M2-refleksen kræver informationsbehandling i hjernen. De sensoriske informationer fra en muskel går op forbi hjernen og tilbage igen for at aktivere de motoriske neuroner. Fordi refleksen skal en længere vej gennem nervesystemet, tager den også længere tid, ligesom det er sandsynligt at der er en eller anden form for informationsbehandling i de højere dele af centralnervesystemet i forbindelse med refleksen.
‘Triggered reactions’ er når musklen reagerer på et stimuli, der ikke kommer fra musklen, men fra andre sanser. Et eksempel kan være bøjerefleksen i menneskets gangcyklus, hvis vi træder på noget spidst. En refleks vi ganske vist kan undertrykke, men som er ubevidst.
Den sidste refleks er den generelle reaktionstid på sensorisk feedback. Et eksempel kan være når en angrebsspiller i basketball laver en finte. Forsvarsspilleren reagerer på angrebsspillerens første bevægelse, og inden forsvarsspilleren opfatter, at det var en finte, går der et lille stykke tid. Den generelle reaktionstid er omkring 120-180 ms. Dette fænomen betyder også, at en finte kan laves så hurtigt, at forsvarsspilleren ikke når at blive snydt.
Sat i relation til det motoriske hierarki, er det sandsynligt at M2-reflekserne foregår på muskelgruppeniveau (niveau 3), da M2-refleksen er den hurtigste, der kan påvirkes ved læring, og muskelgruppeniveauet er det laveste niveau for korrektioner på basis af feedback. Triggered reactions foregår formentlig også på muskelgruppeniveau, mens den generelle reaktionstid formentlig går over flere niveauer, og derfor tager længere tid (Agerskov, 1994).
Ved langsommere færdigheder, kan man måske nå at lave minimale justeringer undervejs, mens det er umuligt ved hurtigere færdigheder, der udføres hurtigt. Før feedback har nået at give besked om bevægelsens aktuelle tilstand, er det for sent at korrigere den hurtige bevægelse. Ved de rigtig hurtige bevægelser, når M1 og M2 tilbagemeldingen ikke engang rundt i feedbackloopet. Hvis en hurtig bevægelse først er sat i gang, kan man ikke bare lave den om igen. Et godt eksempel er en spiller, der er ved sparke på mål i fodbold. M2-refleksen har sendt beskeder fra muskeltenerne om at hoftemuskulaturen er strakt ud over sin normallængde, og at den skal gøre sig klar til at trække sig sammen. Proprioceptorerne i foden sender med en triggered response-refleks besked til fodens stabilisatormuskler om at stramme lidt op, fordi underlaget er ujævnt. I samme øjeblik råber en medspiller på bolden, men han afslutter alligevel selv. Medspilleren kan ikke forstå, hvorfor han ikke bare stoppede benet, mens afslutteren ikke kan forstå, hvorfor medspilleren ikke kunne se, at han var i gang med at afslutte. Han har en fornemmelse af, at han ikke kunne stoppe benet, og han har formentlig ret.
Det ses også i tabel 1, at reaktionstiden stiger, hvis udøveren modtager instruktioner undervejs, eller hvis antallet af valgmuligheder øges. I tabellen ses også, at der går mindst 80-180 ms inden mennesket kan reagere på et stimuli fra en ekstern kilde. I konkurrencesituationer ser vi at badmintonspillere når at vinkle slagene i forhold til modstanderens bevægelser. Dette er sandsynligvis kun muligt for øvede spillere, og det er sandsynligt at denne anticipation foregår i en tæt knytning mellem sansningen og organniveau 4 eller 5, så de visuelle informationer ikke behøver at blive behandlet kognitivt.
Det har også den implikation, at den komplicerede, ballistiske færdighed allerede overstået, inden individet kan nå at processere verbal feedback, som skal behandles på niveau 6. Feedback er altså noget underviseren må give, når udførelsen af færdigheden er overstået.
Hvis færdigheden tager mere end 300 ms, kan kroppen formentlig nå at få tilbagemelding fra en ekstern kilde om bevægelsens aktuelle tilstand, så bevægelsen måske kan nå at blive korrigeret undervejs. Men det betyder stadig ikke, at vedkommende kan tage mod verbal feedback, mens færdigheden udføres, idet det tager et stykke tid fra perceptionen af feedback til udførelsen og inkorporering af feedback. Feedback fra en underviser under udførelsen af færdigheden kræver formentlig bevægelser, der er langsommere end 1 sekund eller cykliske af natur.
4 Opsamling og konklusion
Denne artikel opstiller følgende forståelsesramme for motorisk kontrol. En færdighed er repræsenteret i hjernen som et topologisk kort. Systemet er selvorganiserende gennem det motoriske hierarki og konfigurerer sig selv forskelligt fra gang til gang ud fra en sum af alle påvirkninger på systemet, målet med opgaven og de eksterne kræfter. Den topologiske repræsentation er suppleret af en kinæstetisk repræsentation, der gemmer fornemmelsen af bevægelsen. Disse repræsentationer betegnes som skemaer.
Det motoriske skema er en aktiv organisation, der fastholder det stabile og gentagne fra situation til situation og samtidig er åben for variable faktorer. Hierarkiet er den struktur, der besidder disse egenskaber. Det motoriske hierarki er nervesystemets anticipation af, hvad der kræves for at udføre en færdighed - og denne anticipation virker helt ned på muskelgruppeniveau. Derved bliver hierarkiet et udtryk for nervesystemets erfaringer. De parametre hierarkiet fungerer under udtrykker skemaets åbenhed, og den selektive potentiering udtrykker, hvad nervesystemet forventer kan ske - og derved forbereder sig på (Agerskov, 1994). Kun på niveau 6 er vi bevidste om, hvad der sker. Det er niveauet for taktik og refleksion, som vi ofte godt kan forklare. Vi kalder kroppens viden tavs, fordi den i selve bevægelserne organiserer sig selv gennem det motoriske hierarki.
Teorien beskæftiger sig hovedsageligt med, hvordan en færdighed kontrolleres i udførelsesfasen, og derved udelukkes de psykologiske aspekter i planlægningsfasen. Disse aspekter kan dog inddrages i den fremlagte teori via emotionelle repræsentationer. Bernsteins teori forudsiger et todelt kontrolsystem; et hukommelsesspor, som sætter færdigheden i gang, og et andet som skal forsøge at være med til at rette fejlene. Når man udfører en færdighed, har man altså et system som sørger for at sammenligne den aktuelle bevægelser med de forventede bevægelser.
Denne opfattelse af motorisk kontrol har den fordel, at den kan beskrive den fleksible måde vi bruger kroppen på, og den kan samtidig favne opfattelsen af, at koordination er en temporal samling af muskelkomplekser, som skal løse et bestemt bevægelsesproblem på et givent tidspunkt (Whiting, 1992), og samtidig skaber den plads til, at der kan ske læring i systemet.
Denne forståelse kan også tage højde for eksterne kræfter og også være med til at forklare, hvorfor en udøver pludselig kan have vanskeligt ved at klare en færdighed, som vedkommende tidligere har mestret, ligesom teorien kan give en plausibel forklaring på, hvor mental træning kan virke i forbindelse med motoriske færdigheder. Et grundelement i mental træning er jo netop, at man ikke kan forbedre ens præstation af en færdighed uden at have nogen forkundskaber. Derved kan man teoretisk set tænke sig, at man kan styrke det topologiske og kinæstetiske skema ved mental træning (Agerskov, 1994). Forståelsen kan også forklare, hvordan en spiller pludselig i en presset situation kan præstere en løsning på et motorisk problem, som egentlig ikke burde kunne lade sig gøre (de såkaldte kampslag).
Den fremlagte model for motorisk kontrol og læring er efter min vurdering teoretisk robust. Teorien rejser dog stadig nogle ubesvarede spørgsmål, bl.a.: Hvor er repræsentationerne lokaliseret? Hvordan kan en hjerne, der fungerer vha. fysiologiske processer tænke i helheder? Teorien om motorisk læring har det udgangspunkt, at færdighedstilegnelse foregår gennem små forbedringer, men det behøver ikke altid at være tilfældet (Dickinson, 2004). Fordi den fremlagte forståelsesramme tænker den motoriske repræsentation som en helhed, åbnes der imidlertid mulighed for springvise forbedringer. Teoriens største udfordringer er den empiriske understøttelse. Jeg har imidlertid ikke kunnet finde studier, der kan afvise præmisserne i teorien, så den kan være et godt udgangspunkt for yderligere drøftelse af motorisk kontrol.
Med en viden om hvordan motoriske færdigheder er kontrolleret har man som træner et godt udgangspunkt for at undervise. Det næste spørgsmål man bør stille sig selv handler om læring. Hvor motorisk kontrol er et spørgsmål om organiseringen af bevægeapparatet her-og-nu, så er motorisk læring et spørgsmål om, hvordan sådan en organisering overhovedet bliver mulig og optimeres. Motorisk læring er transformationsprocesser, der relativt permanent ændrer individets præstationspotentiale (Christina, 1997; Schmidt, 2000; Magill, 2001). Hvordan kan man optimere den intern proces der skaber og genskaber færdighederne i udøveren, når vedkommende øver sig? Hvilken rolle kan underviseren spille i læringsprocessen? Hvordan bør feedback udformes for at være mest effektivt? Jeg vender tilbage til nogle af de spørgsmål i senere artikler.
Referencer
Skill Acquisition in Sport - Research, Theory and Practice
Skill Acquisition in Sport gives academics, students, coaches and practitioners the broadest and most scientifically rigorous grounding in the principles and practice of the field. Fully revised, updated and restructured, the third edition integrates theory and practice, and provides more material on practical application than ever before.
Motor Learning and Performance - From Principles to Application
Written in an accessible style, Motor Learning and Performance, 6th Edition enables students to appreciate high-level skilled activity and understand how such performances happen. The text builds a conceptual model of factors that can influence motor performance. As well as, outlining how motor skills are acquired and retained with practice. This brand new sixth edition has been revised to incorporate the most important research findings in the field.
Motor Learning and Control: Concepts and Applications
En af biblerne inden for motorisk læring og kontrol, som er løbende bliver revideret for at være up to date.
[Abernethy 1992] Abernethy, Bruce AND Sparrow, W.A., “The Rise and Fall of Dominant Paradigms in Motor Behaviour Research”, in Approaches to the Study of Motor Control And Learning (Elsevier Science Publishers, 1992).
[Adams 1987] Adams, Jack A., “Historical review and appraisal of research on the learning, retention, and transfer of human motor skills”, Psychological Review 101 (1987), pp. 41-74.
[Agerskov 1994] Agerskov, Henrik, Mental træning og motorisk kontrol (Københavns Universitet. Idræt., 1994).
[Arbib 1989] Arbib, Michael A., The Metaphorical Brain 2. Neural Networks and Beyond (John Wiley and Sons, 1989).
[Bernstein 1984a] Bernstein, N., “Some emergent problems of the regulation of motor acts”, in Human Motor Actions. Bernstein Reassessed (North-Holland, 1984), pp. 343-372.
[Bernstein 1984b] Bernstein, N., “The problem of the interrelation of co-ordination and localization”, in Human Motor Actions. Bernstein Reassessed. (North-Holland, 1984), pp. 77-170.
[Christina 1997] Christina, Robert W., “Concerns and Issues in Studying and Assessing Motor Learning”, Measurement in Physical Education and Exercise Science I, 1 (1997), pp. 19-38.
[Crow 1998] Crow, J. L., “The neural control of human movement”, in Human Movement (Churchill Livingstone, 1998).
[Dickinson 2004] Dickinson, John AND Weeks, David, “One-trial motor learning”, in Skill Acquisition (Routledge, 2004), pp. 63-83.
[Feigenberg 1998] Feigenberg, Josef M., “The Model of the Future in Motor Control”, in Progress in Motor Control. Volume One. Bernstein’s tradtions in Movement Studies vol. 1, (Human Kinetics, 1998).
[Gallistel 1980] Gallistel, C. R., The Organisation of Action: a new Synthesis (Hillsdale, New Jersey: Lawrence Erlbaum Associates Publishers, 1980).
[Gurfinkel 1998] Gurfinkel, Victor S. AND Cordo, Paul J., “The Scientific Legacy of Nikolai Bernstein”, in Progress in Motor Control. Bernstein’s traditions in movement studies (Human Kinetics, 1998).
[Hodges 2004] Hodges, Nicola J. AND Franks, Ian M., “Instructions, demonstrations and the learning process”, in Skill Acquisition in Sport (Routledge, 2004), pp. 145-174.
[Latash 1998] Latash, Lev P., “Automation of Movements: Challenges to the Notions of the Orienting Reaction and Memory”, in Progress in Motor Control. Berntstein’s Tradtions in Movement Studies (Human Kinetics, 1998).
[Lüders 2001] Lüders, Kurt, “Hvorfor tidlig specialisering?”, Focus 2 (2001), pp. 70-74.
[Magill 2001] Magill, Richard A., Motor Learning. Concepts and Applications (McGraw Hill, 2001).
[Newell 1984] Newell, Karl M., “Coordination, Control and Skill”, in Differing Perspectives on Motor Learning, Memory, and Control (1984).
[Schmidt 1975] Schmidt, Richard A., “A schema tehory of discrete motor skill learning”, Psychological Review 82 (1975), pp. 225-260.
[Schmidt 1997] Schmidt, Richard A., “Continuous Concurrent Feedback Degrades Skill Learning: Implications for Training and Simulation”, Human Factors 39, 4 (1997), pp. 509-525.
[Schmidt 2000] Schmidt, Richard A. AND Wrisberg, Craig, Motor Learning and Performance. A Problem-based Learning Approach (Human Kinetics, 2000).
[Schmidt 2003] Schmidt, Richard A., “Motor Schema Theory After 27 Years: Reflections and Implications for a New Theory”, Research Quarterly for Exercise and Sport 74, 4 (2003), pp. 366-375.
[Selch 2003] Selch, Henrik AND Østergaard, Jan, Powerfodbold (Forlaget Goldfingers, 2003).
[Singer 1990] Singer, Robert N., “Motor Learning Research: Meaningful for Physical Educators or A Waste of Time?”, Quest 42 (1990), pp. 114-125.
[Summers 2004] Summers, Jeffrey J., “Historical perspective on skill acquisition”, in Skill Acquisition in Sport (Routledge, 2004), pp. 1-26.
[Weisendanger 1998] Weisendanger, Mario, “Bernstein’s principle of Equal Simplicity”, in Progress in Motor Control. Volume One. Bernstein’s tradtions in Movement Studies (Humans Kinetics, 1998).
[Whiting 1992] Whiting, H. T. A. AND Vogt, Beatrix, “Human Skill and Motor Control: Some Aspects of the Motor Control - Motor Learning Relation”, in (Elsevier Science Publisers, 1992).
You can be the first to add a comment below.
Leave a Comment
Din e-mail bliver ikke offentliggjort. Obligatoriske felter er markeret Required fields are marked *.